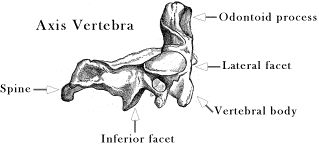
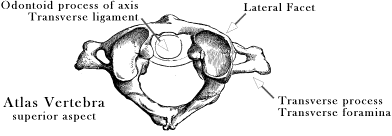
Spinal Dynamics I: The Axio-atlanto-occipital Assemblage Bones interact through joints. The relative placements of bones across joints determine how they move in space. In this section we will consider two related assemblages of bones. The first is the atlanto-occipital and atlanto-axial joints, which together marry the skull to the vertebral spine. The movements in the axio-atlanto-occipital assemblage are complex because of the interactions between these bony elements (Kapandji 1974; Hoppenfeld 1976; White and Panjabi 1978; Dvorak and Panjabi 1987; Dvorak, Penning et al. 1988; Dvorak, Schneider et al. 1988; Nordin and Frankel 1989; Panjabi and White 1989; Penning 1989; White and Panjabi 1990; Ghanayem, Zdeblick et al. 1998; Panjabi, Dvorak et al. 1998; Bogduk 1999; Bogduk and Mercer 2000; Levangie and Norkin 2001; Standring 2005; Langer 2005m; Langer 2005p). The movements in this joint complex are unlike those in the rest of the spine, both in type and magnitude of angular excursion. In related matters, the vertebral artery passes between the transverse processes of the atlas and the axis, which places it far from the center of rotation for those two bones, and the amount of rotation between those bones in much greater than between any other vertebrae. Both of these anatomical factors place the segment of the vertebral artery between the two bones at substantial risk of excessive strain, making it the part of the artery most apt to be injured by physical activity or therapeutic manipulation. We will briefly consider this anatomy and its implications for blood flow as part if our consideration of the axio-atlanto-occipital assemblage. Blood flow is considered in considerably greater detail elsewhere (Langer 2005m; Langer 2005n; Langer 2005o; The second bony assemblage is the remainder of the cervical spine, the lower cervical spine. The lower cervical spine is one of the more flexible sections of the spinal column and it moves about an interesting type of facet joint that allows movements that do not fit easily into any of the cardinal planes (Kapandji 1974; White and Panjabi 1978; Nordin and Frankel 1989; Panjabi and White 1989; Penning 1989; White and Panjabi 1990; Milne 1991; Committee 1998; Ghanayem, Zdeblick et al. 1998; Onan, Heggeness et al. 1998; Panjabi, Dvorak et al. 1998; Bogduk 1999; Bogduk and Mercer 2000; Levangie and Norkin 2001; Standring 2005; Langer 2005q; Langer 2005r; Langer 2005s). The facet joints of the second cervical vertebra,C2, to the first thoracic vertebra, T1, are inclined at about a 45° angle to the coronal plane of the vertebra. As a result, the movements of the cervical spine involve a combination of sideflexion and lateral rotation. Also, since the successive vertebrae are inclined relative to each other, this movement has different consequences for the movements of the cervical spine at multiple levels. The lower cervical spine provides an excellent opportunity to model an assemblage of bones and joints, because it is essentially the same element repeated several times, which greatly reduces the numbers of parameters that need to be manipulated. The Axio-atlanto-occipital AssemblageAnatomy: The inferior aspect of the second cervical vertebra, the axis, is like the other cervical vertebrae in having inferior facets that face anteriorly, inclined about 45° to the coronal plane and an intervertebral disc that joins two vertebrae in an uncinate (U-shaped) joint. The upper surface of the axis is quite different. We will deal with that type of joint in some detail, below. The rest of the bone is of more interest at this point. The axis has a long, tooth-like, odontoid process, or dens, that extends rostrally, through the ring-like anterior arch of the atlas, the first cervical vertebra. The odontoid process is the embryological remnant of the vertebral body of the atlas, which has become fused to the vertebral body of the axis. It is held in place by a transverse ligament that passes posterior to it, joining the two sides of the atlas and completing a tubular socket that encloses the peg-like odontoid process. The axis and atlas vertebrae abut upon each other in a pair of lateral facet joints that form the lateral wall of the tube and allow and support lateral rotation between the two vertebrae. The lateral facets are tilted posteriorly and laterally, so that the support surface is slightly inclined. The odontoid process provides a fulcrum for lateral rotation of the atlas upon the axis. Consequently, the axis of rotation for the atlanto-axial joint lies within the odontoid process. During lateral rotation, the lateral facets slide upon each other. The surfaces of the facets in this joint are almost flat, only slightly convex in their facing surfaces. The modest curvature of both facet surfaces allows the two bones move a bit closer as they side over each other, allowing a small amount of additional rotation. The overall movement between the two bones is rather like stacked funnels, where the upper funnel rotates about the spout of the lower funnel.
There is also a small amount of play in the odontoid part of the atlanto-axial joint so that the atlas may tilt forward and back through about 10° of sagittal movement. That is a comparatively small amount of movement when compared to the full range of sagittal movement is the cervical spine, but comparable to amount of rotation in most individual intervertebral joints. The principal movement of the atlas about the odontoid process of the axis is lateral rotation. It is estimated that the average range of movement is a bit more than 40°, varying between about 30° and 55° in different individuals, which means about 45° in either direction, for a total of about 90° of lateral rotation in the joint. That is a remarkable amount of movement in a single vertebral joint and it is physiologically important in that the vertebral artery passes just lateral to the lateral articular processes, so it is stretched between the transverse foraminae of the two vertebrae. Consequently, the most common site for mechanical trauma to the vertebral artery is in the segment that passes between C2 and C1.
The joint between the atlas and the occiput, the atlanto-occipital joint, occurs between the lateral articular processes of the atlas and the occipital condyles. The surfaces of those facets are convex inferiorly, so that the occipital condyles effectively sit in a shallow bowl formed by the lateral facets of the atlas. The axis of rotation is transverse and located slightly rostral to the joint, passing through the occiput. The two joint surfaces converge anteriorly, which causes the joint to become tighter as the head tilts into flexion or extension. The range of motion for sagittal movements is estimated to be between 10° and 30°, with the average about 20° of angular excursion. There may a small amount of sideflexion, about 3°, and lateral rotation, about 5.5°, however these may be more joint play than a useful physiological movement. However, the small amount of sideflexion turns out to have physiological implications for the amount of sagittal rotation in the atlanto-axial joint (see below).
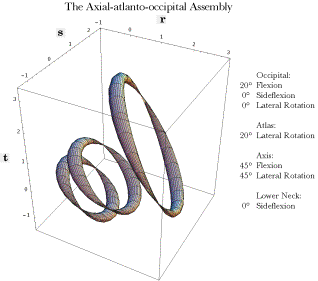
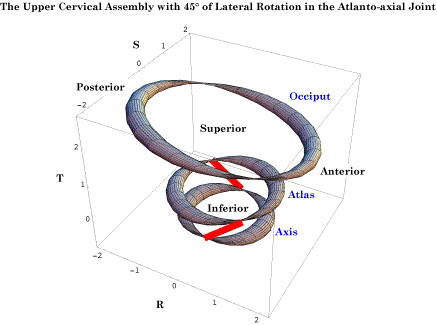
The rings are, from above down, the occiput, the atlas and the axis. The lateral processes of the vertebrae are indicated by the pinched regions of their rings. The occipital ring is at the level of the axis of rotation and the pinched region is the location of that axis. All the bones are in neutral position. The r axis is anterior/posterior and the s axis is medial/lateral. The scale is arbitrary, related to the physical dimensions of the atlas. Modeling the Anatomy with Quaternions: In the following discussion, based on that model, the atlas and axis vertebrae and the occiput have been represented by simplified images, pinched tori, which capture their locations and orientations without obscuring their spatial relationships. Given the rotation in each direction for each element of the joint, the model computes the arrangement of the bones. In neutral position, the three bones are aligned with the lateral processes of the cervical vertebrae in the same coronal plane and the transverse foramina of the atlas lateral to that for the axis. The pinched regions of the rings occur at the locations of the transverse foraminae. The anterior aspects of the vertebrae are to the right. The occipital ring is placed so that the axis of rotation is through the pinched parts of the ring and the ring is horizontal in neutral position. The ring would sit some distance into the skull, rather than at the level of the occipital condyles. In the following figure, rotations have occurred in several of the possible axes of rotation. Rotations occur both within the assemblage and outside it. The lower neck is assumed to have caused the axis to be tilted 45° anteriorly and laterally rotated 45° to the left. The axis is laterally rotated 20° to the left and the occiput is anteriorly tilted 20°. Any combination of rotations may be examined in the model, making it a tool to study the movements in the upper cervical spine.
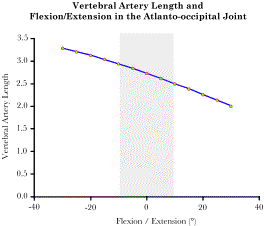
The placements of the occiput, atlas, and axis after the indicated movements about several axes of rotation The Effect of Movements Upon the Intracranial Segment of the Vertebral Artery: Once one has a model of this sort, it is possible to perform calculations that address anatomical questions. For instance, the vertebral artery passes behind the facet joints for the atlanto-occipital joint and enters the vertebral canal to run rostrally through the foramina magnum to the pons in the brainstem, where it fuses with the opposite vertebral artery, to form the basilar artery. The model allows one to determine how flexion and extension of the occiput upon the atlas affects the length of the vertebral artery required to bridge that gap. An extension vector attached, to the occiput, was created to encode the location of the pontomedullary junction within the skull. Another extension vector, attached to the atlas, encodes the location of the vertebral artery as it passes behind the articular facets and is apparently firmly attached to the bone. By examining the distance between those two locations as flexion and extension in the atlanto-occipital joint are varied, one can obtain a fair estimate of the length of the vertebral artery that is necessary to accommodate for the movement. It turns out that the placement of the artery adjacent to the facet joint minimizes the change in distance between the end points of the artery. It is estimated that the range of motion is less than 20° for both directions combined, so, the change in the gap between the extremes of the physiological sagittal movements is about 10% of the gap in neutral alignment. Given that the artery is not normally taut, that is a comparatively small change in distance and we would not expect there to be significant strain in that segment of the artery.
The gap between the lateral facets of the atlas and the fusion of the vertebral arteries has been computed for a range of flexion and extension. The shaded interval corresponds to actual physiological values. The Effects of Movements Between C1 and C2 upon the Vertebral Artery: We can ask a similar question about the distance between the transverse processes of the first and second vertebrae. The question is addressed by creating extension vectors for the transverse foraminae of the two vertebrae. Since the amount of rotations between the vertebrae is considerable, about 45° in each direction, and the artery is located far from the axis of rotation, there is considerable potential for strain in the artery.
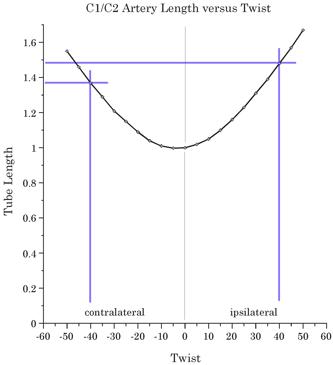
The red line segments indicate the locations of the vertebral arteries between the atlas and the axis when there is a rotation between the two bones. When the calculations are performed it can be seen that there is a substantial increase in the distance that the vertebral artery must span as the rotation occurs. As shown in the following figure, the distance between the transverse process increases about 1.35 to 1.5 times as the neck approaches full physiological lateral rotation. That is more than the artery is able to stretch without tearing. However, arteriograms of the arteries in many persons indicate that there is normally substantial laxity in the artery when the neck is in neutral position, so that the rotation simply takes up the slack instead of straining the artery. Still, the most common site for traumatic damage to the vertebral artery is in the interval between the atlas and the axis. Should the neck be rotated more than it normally does, there is the potential to strain the artery to the point where it is torn. That may become a consideration when a manipulation of the neck is done that suddenly increases the range of motion in the atlanto-axial joint, if the length of the artery had adjusted to a shortened maximal gap. Rotating the head to the endrange of lateral rotation is sufficient in some individuals to cause a cessation of blood flow in a vertebral artery (Arnold, Bourassa et al. 2003; Arnold, Bourassa et al. 2005;
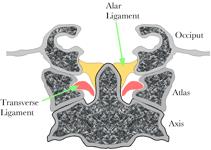
The length of the gap spanned by the vertebral artery is plotted versus the amount of rotation in the atlanto-axial joint. Normal range is about 40° in both directions, but it may be as much as 60° in some individuals. The Role of the Alar Ligaments in Movements of the Upper Cervical Spine: Another situation where the gap may increase substantially is when the lateral rotation is increased by changing the usual physiological constraints upon the movement ( The atlas and axis are joined to the base of the occiput by several fascial membranes and ligaments, which enclose the brainstem and upper spinal cord and lend stability to the region. However, one of these, the alar ligaments, is thought to play a part in restricting the amount of lateral rotation between the axis and the skull. The alar ligaments extend from the inner rim of the foramen magnum, just medial to the occipital condyles, to the apex of the odontoid process of the axis vertebra. Because the atlas lies between those two bones, the alar ligament must affect it and we expect to see consequences both for lateral rotation, which occurs largely between the atlas and the axis, and for sagittal head movements, which occur largely between the atlas and the occiput.
The alar ligaments bind the occiput to the odontoid process of the axis. Consequently it is able to influence flexion/extension and lateral rotations. The alar ligaments are longer than the gap between its bony attachments, therefore it is possible to move the attachment sites for some distance before one or both ligaments become taut and stop the movement. Flexion and extension move the occipital insertion anterior and posterior relative to the odontoid process and therefore one would expect their range to be restricted by the tethering that occurs when the gap between the two insertions becomes equal to the length of the ligament. Similarly, lateral rotation between the occiput and the axis will move the attachment on the odontoid process away from the attachment on the occiput.
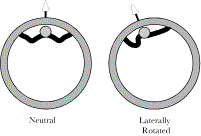
Since the center of rotation for the atlas upon the axis is through the odontoid process, a lateral rotation will move the occipital insertion away from the odontoid insertion on the side opposite the direction of the rotation and move the two attachments closer together on the opposite side. Consequently, the alar ligaments are checks on lateral rotation.
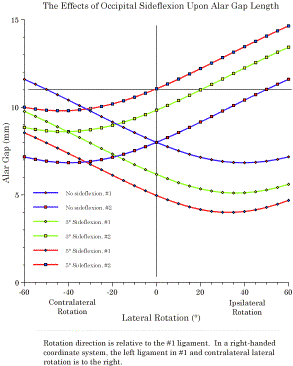
When there is no sideflexion lateral rotations are symmetrical and the alar ligaments appear to be the principal restraint on rotation. With small amounts of sideflexion, ipsilateral rotation is substantially reduced but the alar ligaments cease to be a restraint on contralateral rotation. The horizontal gray line indicated the usual length of an alar ligament. Since we know the relative distances between significant structures in the upper cervical assemblage and the approximate length of the alar ligaments, it is possible to enter the values into the model of the upper cervical assemblage and to examine the implications for movements between the bones in the assemblage. That has been done and the details are available elsewhere (Langer 2005p). We will examine the results of one set of calculations in which the distance between the insertion sites for the alar ligaments are computed for a range of lateral rotation, assuming that the head started in neutral position (the blue lines), sideflexed 3° (green lines), or sideflexed 5° (red lines). The gap for each ligament is computed. When there is no sideflexion, the relationships are symmetrical (blue lines) and the head can rotate about 40-45° before it is stopped by a taut alar ligament. That is about the range of lateral rotation that is normally observed. If the head is sideflexed (green and red lines), then the relationship is asymmetrical and shifted so that there is less ipsilateral rotation before the ligament becomes taut, but more contralateral rotation. In fact the alar ligament effectively ceases to be a restriction upon lateral rotation to the contralateral side. Other ligaments must come into play, because at a point not that much greater than 45° of lateral rotation the lateral facets will fail to abut and there is a possibility of them subluxating and locking in a side-by-side position. There is also a risk of impingement of the vertebral arch upon the cervical spinal cord. Occasionally, individuals tear one alar ligament and it is observed that they gain a substantial amount of lateral rotation, but the rotation is stopped short of catastrophic impingement, although their facet joints can become locked in endrange lateral rotation. The model of the upper cervical assemblage predicts the approximate range of lateral rotation without sideflexion and it indicates that the increased range when the head is sideflexed may well be due to the reduction of the gap that occurs with sideflexion. Not considered here is the observation that the model predicts that when the head is flexed or extended in the atlanto-occipital joint, then there is a substantial reduction in the amount of lateral rotation that can occur before the alar ligaments become taut and stop the movement (Langer 2005p).
Arnold, C., R. Bourassa, et al. (2003). "Doppler studies evaluating the effect of a physical therapy screening protocol on vertebral artery blood flow." Manual Therapy 9/1: 13-21. Bogduk, N. (1999). "The neck." Baillieres Best Pract Res Clin Rheumatol 13 Bogduk, N. and S. Mercer (2000). "Biomechanics of the cervical spine. I: Normal kinematics." Clin Biomech (Bristol, Avon) 15 Committee, T. C. S. R. S. E. (1998). The Cervical Spine. Phiadelphia, Lippincott - Raven. Dvorak, J. and M. M. Panjabi (1987). "Functional anatomy of the alar ligaments." Spine 12: 183-190. Dvorak, J., L. Penning, et al. (1988). "Functional diagnostics of the cervical spine using computer tomography." Neuroradiology 30 Dvorak, J., E. Schneider, et al. (1988). "Biomechanics of the craniocervical region: the alar and transverse ligaments." J Orthop Res 6 Ghanayem, A. J., T. A. Zdeblick, et al. (1998). Functional Anatomy of Joints, Ligaments, and Discs. The Cervical Spine. T. C. S. R. S. E. Committee. Philidelphia, Lippincott - Raven: 45 - 52. Hoppenfeld, S. (1976). Physical Examination of the Spine and Extremities. New York, Appleton-Century-Crofts. Kapandji, I. A. (1974). The Physiology of the Joints. Annotated diagrams of the mechanics of the human joints. New York, Churchill Livingstone. Levangie, P. K. and C. C. Norkin (2001). Joint Structure and Function. A Comphrehensive Analysis. Philadelphia, F. A. Davis Company. Milne, N. (1991). "The role of zagapophyseal joint orientation and uncinate processes in controlling motion in the cervical spine." J. Anat. 178 Nordin, M. and V. H. Frankel (1989). Basic Biomechanics of the Musculoskeletal System. Philadelphia, Lea & Febiger. Onan, O. A., M. H. Heggeness, et al. (1998). "A motion analysis of the cervical facet joint." Spine 23(4): 430-9. Panjabi, M. M., J. Dvorak, et al. (1998). Cervical Spine Kinematics and Clinical Instability. The Cervical Spine. T. C. S. R. S. E. Committee. Philadelpia, Lippincott - Raven: 53 - 78. Panjabi, M. M. and A. A. White (1989). Biomechanics of Nonacute Cervical Spinal Cord Injuries. The Cervical Spine. T. C. S. R. S. E. Committee. Philadelphia, J.B Lippincott: 91 - 96. Penning, L. (1989). Functional Anatomy of Joints and Discs. The Cervical Spine. T. C. S. R. S. E. Committee. Philadelphia, J.B. Lippincott: Standring, S. (2005). Gray's Anatomy. The Anatomical Basis of Clinical Practice. Edinburgh, Elsevier, Churchill Livingstone. White, A. A. and M. M. Panjabi (1978). Clinical Biomechanics of the Spine. Philadelphia, J. B. Lippincott. White, A. A. and M. M. Panjabi (1990). Clinical Biomechanics of the Spine. Philadelphia, J.B. Lippincott.
|